


Thyroid Hormone and Spotted Coats
Patterning of coat colours—piebaldness—is determined during fetal development


[Adapted from my 2006 book, Rhythms of Life, with selected references]1
Although the size, shape and reproductive changes in biological traits associated with the early stages of domestication have been shown to be under the control of thyroid hormone, the appearance of piebaldness – black or any other solid colour marked with white – is the hardest to explain in this context.
Think spotted cows and horses with white feet, as well as Samoyed dogs and totally white sheep. The high incidence of whitespotting or piebaldness in domestic animals has always been somewhat of an enigma and is usually assumed to be a consequence of deliberate selection.
However, piebald markings are quite rare in most wild mammals.

Piebaldness is also rare in the ancestors (or in close living relatives) of all domestic animals, although it is the norm in a small number of groups (such as skunks, killer whales, and giant panda, among others).
This low natural incidence of piebaldness in wild populations makes the prospect of increasing the proportion of individuals with this trait, via natural or human selection, not just improbable, but almost impossible.
However, the early appearance of piebaldness in the silver foxes experimentally selected for stress-tolerant behaviour (described in detail below) suggests that piebaldness could be an inevitable consequence of the early domestication process.
To explain why this is so, we need a short discussion on how pigment and patterned coats are generated.
Pigment (technically called melanin) is produced in special cells (melanocytes) that exist within skin, hair follicles and other body tissues. Many different factors can affect the activity of pigment cells during skin and hair colour production, including hormonal influences that come from the pituitary (via MSH, melanocyte-stimulating hormone) as well as spontaneous mutations affecting the genes and various cofactors involved.
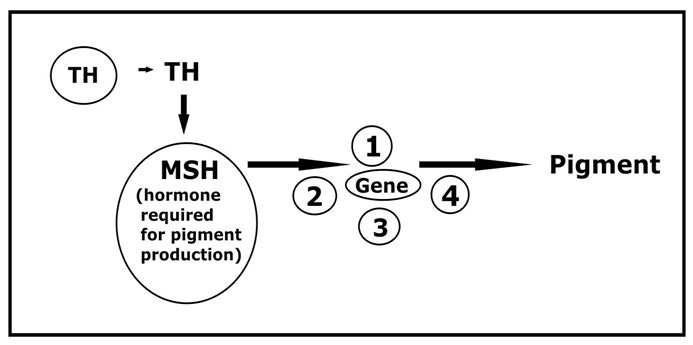
It’s clear that small genetic mutations or varying amounts of hormone (and perhaps both) can result in different coat and skin colours being produced.2
However, patterning of colour—piebaldness—is determined during fetal development.3
The explanation that so far best accounts for colour patterns like whitespotting is that during early embryonic development, pigment cells start from a few locations and move throughout the embryo. Seven pairs of immature pigment cells seem to be required, and these migrate from their origins at the central mid-line of the tiny embryo (called neural crest tissue) to all areas of the body.
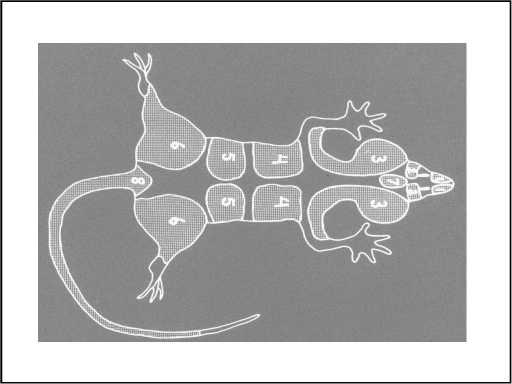
Through controlled migration and multiplication of these immature pigment cells and their subsequent maturation into functional pigment cells, the entire embryo becomes completely pigmented.
Typically, colour appears to spread from two single patches of pigment-producing cells (one at the base of the tail and another between the eyes—numbers 8 and 7, respectively) and from six pairs of symmetrical patches over the hips, lumbar region, rib cage, ears, eyes and nose—fourteen areas in all. Regions in between these centres that do not receive pigment-producing cells remain white, either because cell migration has been delayed or because multiplication of the cell patches prematurely stops.
The result is a coat colour pattern that is variously spotted, like the border collie below.

In a now-famous selection experiment that began in Siberia in the late 1950's, geneticist Dmitri Belyaev began with a large population of silver foxes, a naturally occurring form of the red fox, Vulpes vulpes, that’s black with white-tipped hairs and a white-tipped tail (see photo below). Silver foxes were being farmed commercially for their fur as an alternative to trapping wild animals. These farmed silver foxes retained all of the characteristics of wild foxes, including being timid around people, which meant they often did not reproduce well.4

Belyaev wondered if selecting among these captive foxes for those with less excitable temperaments might generate an animal that was easier to farm commercially.
Belyaev assessed a randomly selected 465 foxes from several fur farms for their response to a gloved hand extended into each animal’s cage, choosing only those showing a quiet, tolerant curiosity (without fear or aggression) for further breeding.
These curious foxes, about 10% of the total, demonstrated noticeably less fearful behaviour toward humans than the others, what I would call stress-tolerant.
Interestingly, when farm breeding records were checked, it was noticed that the females in this less fearful population that had previously given birth had all been early breeders: they had consistently ovulated within the first week or two of the six-to eight-week fox breeding season. This correlation suggested that a pre-existing link existed between the behavioural response to stress (the “gloved-hand intruder”) and the timing of ovulation in these foxes, although the researchers didn’t understand what this might mean.
After several generations of mating stress-tolerant, early breeding females with stress-tolerant males and selecting among each year’s litter of pups for tolerant behaviour only, the time of ovulation (necessary for breeding) as well as the annual moult for many females shifted forward by several months (to October or November, from early February).
As the experiment continued, ovulation and hair moult receded further still, until several females were ovulating twice a year (two full reproductive cycles). After twenty generations (twenty years), some females were able to produce two litters a year.
Completely unexpectedly, however, early on a number of other novel traits suddenly appeared, including a curled tail, flopped-over ears, a white “star” on the forehead, a pattern of distinctive brown markings that the researchers called “brown piebald,” and a classic white piebald pattern of black-and-white markings similar to that seen in a typical Border collie, as shown below from a photo provided in Belyaev’s 1979 paper.

Belyaev described the foxes with these novel physical and physiological traits as having "doglike" behaviour because they barked and were quite unafraid of people—they would even crawl into his lap when he sat in a large enclosure with them.
And although the experiment was a failure as far as the fox farms were concerned – because the farmers certainly did not want black-and-white-spotted foxes – Belyaev recognized that the results were scientifically important and wrote them up for publication.
Follow-up studies on the selected foxes continued for decades – indeed, I visited the experimental farm in 2014. For example, the extreme piebald coat patterns that left foxes looking like a border collie, were ultimately determined by researchers to be caused by a one- to two-day delay in the migration rate of pigment-producing cells in the embryo.5
Genetic analyses confirmed that the changes in the selected foxes were not caused by spontaneous mutations in any known gene, or in fact by any known biological mechanism. Additional experiments undertaken later—by other scientists in other countries, and involving red foxes (in the US) and fallow deer (in Germany)—gave similar kinds of results.
Belyaev had concluded that something in the selection for tolerant behaviour toward humans was not only causing juvenilization of physical form and behaviour, but was somehow “disrupting” the normally constrained timing of sexual reproduction—he just couldn’t explain how or why.
However, the fact that none of the characteristics found among the selected foxes had been present in the original population suggests that all might be inevitable consequences of the selection for stress-tolerant behaviour.
We now know slightly more about this process, but not much.
There are at least three genes which can cause disruption of the normal migration of pigment-producing cells during early development: piebald (S); dominant whitespotting (W), and steel (Sl). Specific mutations in each of these genes result in a distinct piebald pattern of pigmentation that in virtually all cases are associated with particular neurological disorders, including deafness (because inner ear sensory cells also originate in the neural crest).6
Due to the associated neurological defects, experimental laboratory mice with these mutations can’t possibly possess the same genetic makeup as naturally piebald wild animals or most piebald domesticates. It’s pretty clear, for example, that the extreme piebaldness (“one big spot”) that’s the normal colour for most white domestic and wild animals that have black-eyes (such as the Samoyed dog and the polar bear) is not associated with the lethal digestive system defects or the deafness linked with the mutant genes in all-white experimental animals.
However, there is a strong link between early movement of neural crest cells and thyroid hormone.
Both T4 and retinoic acid (a vitamin A derivative) have been identified as essential to the orderly movement of cells out of the neural crest during early development. The T4 needed for neural crest cell migration is required by the embryo very early in development and, in mammals (including humans), the maternal thyroid gland has been shown to be the source of the T4 utilized.7
Still, a direct correlation between disruption of thyroxine production and piebaldness has not yet been demonstrated but the circumstantial evidence in favour of such a relationship is very strong.
Obviously, piebald coat patterns can result from several distinct abnormalities, not all of which are genetic. Unfortunately, no detailed scientific studies appear to have been done on the specific types of piebaldness possessed by wild animals, and the role of thyroid hormone in this process has not been specifically investigated.
However, because piebald coat colour patterns are now known to reflect changes in embryonic growth, their consistent appearance in domestic animals suggests that piebaldness may be an incidental consequence of the changes in growth rates known to be associated with the early domestication process—and this may be true in many wild animals as well.
More research definitely needs to be done on this topic, and there is a lot we will never know. For example, although piebald markings do occur in modern grizzly populations (usually a white chest or collar), it’s unknown whether the ancient grizzly populations that polar bears descended from were similarly marked.8
Crockford, S.J. (2006). Rhythms of Life: Thyroid Hormone and the Origin of Species. Trafford, Victoria.
Hearing, V.J. (1993). Invited editorial: Unraveling the melanocyte. American Journal of Human Genetics 52, 1-7.
Searle, A.G. (1968). Comparative Genetics of Coat Colour in Mammals. Logos Press Ltd., London.
Silvers, W.K. (1961). Genes and the pigment cells of mammals. Science 134, 368-373.
Silvers, W.K. (1979). The Coat Colors of Mice: A Model for Mammalian Gene Action and Interaction. Springer-Verlag, New York.
Belyaev, D.K. (1979). Destabilizing selection as a factor in domestication. Journal of Heredity 70, 301-308. https://doi.org/10.1093/oxfordjournals.jhered.a109263
Prasolova, L.A., Trut, L.N. and Vsevolodov, E.B. (1997). Morphology of mottling hairs in domesticated silver foxes (Vulpes vulpes) and relation between the expression of the star and the mottling mutation, Evolutionary-genetic and Genetic-physiological Aspects of Fur Animal Domestication: A Collection of Reports, L.N. Trut and L.V. Osadchuk (eds.), IFASA/Scientifur, Oslo, p. 31-40.
Belyaev, D.K., Ruvinsky, A.O. and Trut, L.N. (1981). Inherited activation-inactivation of the star gene in foxes: its bearing on the problem of domestication. Journal of Heredity 72, 267-274.
Belyaev, D.K., Trut, L.N. and Ruvinsky, A.O. (1975). Genetics of the W locus in foxes and expression of its lethal effects. Journal of Heredity 66, 331-338.
Bennett, D.C. (1991). Color genes, oncogenes and melanocyte differentiation. Journal of Cell Science 98, 135-139.
Hosoda, K., Hammer, R.E., Richardson, J.A., et al. (1994). Targeted and natural (piebald lethal) mutations of endothelin-B receptor gene produce megacolon associated with spotted coat color in mice. Cell 79, 1267-1276.
Little, C.C. (1958). Coat color genes in rodents and carnivores. Quarterly Review of Biology 33, 103-137.
Witte, O.N. (1990). Steel locus defines new multipotent growth factor. Cell 63, 5-6.
Barres, B.A., Lazar, M.A. and Raff, M.C. (1994). A novel role for thyroid hormone, glucocorticoids and retinoic acid in timing oligodendrocyte development. Development 120, 1097-1108.
Chan, S. and Kilby, M.D. (2000). Review: Thyroid hormone and central nervous system development. Journal of Endocrinology 165, 1-8. https://joe.bioscientifica.com/view/journals/joe/165/1/1.xml
Cowling, K., Robbins, R.J., Haigh, G.R., et al. (1994). Coat color genetics of Peromyscus: IV. Variable white, a new dominant mutation in the deer mouse. Journal of Heredity 85, 48-52.
Crockford, S.J. (2023). Polar Bear Evolution: A Model for How New Species Arise. Amazon KDP, Victoria.