
Island Syndrome Part 1: Introduction
The thyroid phenotype of founders explains dwarfs, giants, and flightless birds on islands

[An audio version of Parts 1 and 2 are available in Episode 16 of my podcast, Bites Out Loud, here]

Island syndrome is the biological term used to describe the tendency for unique island vertebrates to possess different body size and limb proportions compared to mainland ancestors. Although a number of suggestions have been offered to explain this phenomenon of altered growth rates on islands, no formula given so far fits all situations. I contend that the individual thyroid hormone phenotype of mainland founders determines the life history traits and body size of island descendants: it matters which individuals come first.
[Adapted and updated from my 2006 book, Rhythms of Life, with selected references, including some updated ones]1
Most animal populations on islands show similar trends over time: small mammals get larger (as do some birds), large mammals get smaller (as do some small animals, like rabbits and shrews), and birds often become flightless with enlarged bills and well-developed hind limbs. The graphic below illustrates this tendency.
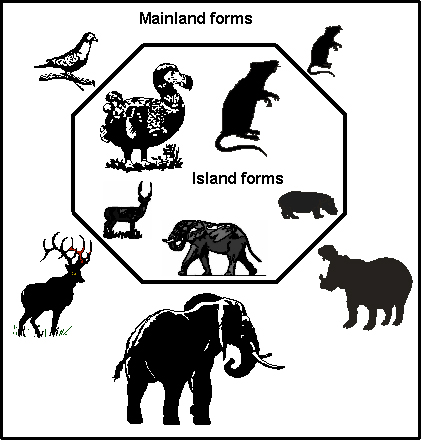
Some of the unique animal species found on remote and continental islands, called endemic species, can be bizarrely different from their mainland counterparts. Sometimes, island-dwelling animals are so different in size or shape that determining their closest relatives was a real challenge before DNA technology came along. Many are, or were, especially vulnerable to rapid extinction, particularly from the hunting practices peculiar to humans: the recently exterminated dodo of Mauritius and great auk of Newfoundland and Britain are recent examples, while the pygmy hippo of Cyprus likely fared the same fate in prehistoric times.2
The intriguing phenomenon of island endemism has captured popular interest for decades, but most island oddities vanished so long ago they are poorly known outside the scientific world. However, within that specialist realm, island-dwelling animals have been the focus of study for evolutionary biologists and ecologists for more than a century, beginning with Alfred Russell Wallace’s fascinating account of his studies in Indonesia, published back in 1880.
Much of this scientific interest is fuelled by the fact that animal populations on islands develop under such controlled conditions they come close to qualifying as natural experiments. As a consequence, in-depth studies of island endemics are particularly well suited to answering questions of how adaptive evolutionary processes actually work. In principle, understanding how these processes work in island situations should help enormously in unravelling how and why giants and dwarfs occur in other circumstances.3
Attention to this topic has peaked again in all quarters in 2004-2005, driven by a contentious debate on whether remains of an extinct hominin from Flores Island in Indonesia should be added to our list of island dwarfs. There have been renewed calls to explain what has become known as island syndrome, the phenomenon of dramatic change in size, shape and behaviour characteristic of isolated island animals as compared to their mainland ancestors.
The specific question that now demands an answer is this: how could one of our closest ancestors (Homo erectus), a species similar in size to ourselves and clever enough to manufacture an arsenal of stone tools, be reduced to a small-brained creature one metre tall (Homo floresiensis) just because it inhabited an island?4
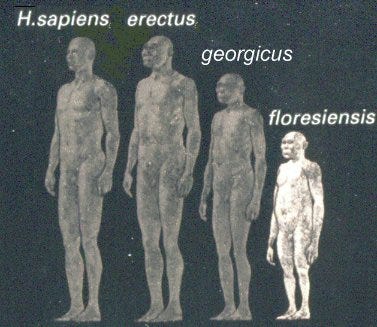
I believe I can answer this question, not just for the hominin curiosities but for all island vertebrates, by applying the concept of thyroid rhythm theory to the phenomenon. Island colonization considered in this light explains not only the parallels that have long been noticed between island endemics and domestic mammals, but also the strong trend toward anomalies of limbs and body shape found among so many island forms. Thyroid rhythm theory provides a more comprehensive explanation for island syndrome than previously proposed hypotheses because it describes a specific biological mechanism to account for these changes that is widely applicable and testable. See my previous essays here, here, and here.
Colonizing Islands
Obviously, remote oceanic islands are difficult to colonize because they’re hard to reach. Small terrestrial mammals and weak flying birds generally end up on oceanic islands by accident (this is often referred to as sweepstakes dispersal). Examples of such accidental colonization include rodents becoming marooned on the vegetative flotsam generated by storms (uprooted trees, etc.) and small forest birds and bats getting blown far out to sea by fierce winds. Migratory birds that are powerful enough to withstand storm winds may nevertheless colonize remote oceanic islands, perhaps out of curiosity or as a result of navigational error.
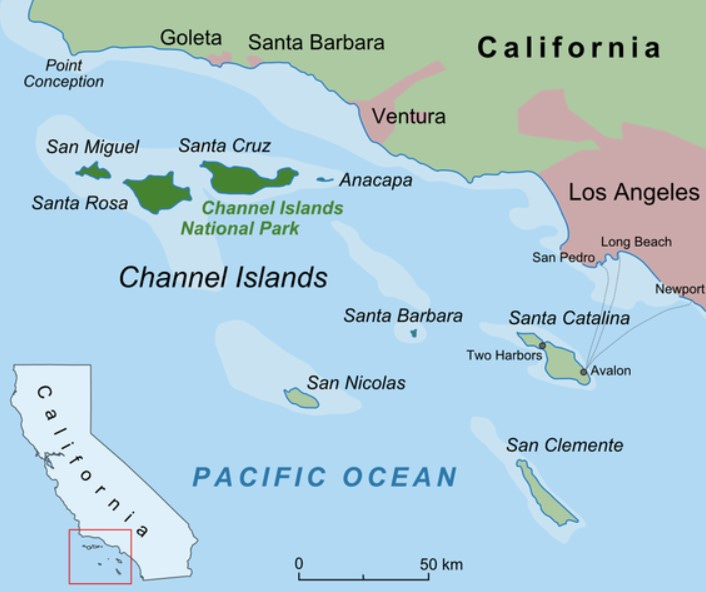
Large mammals are much less likely to colonize isolated oceanic islands, quite simply because they’re too far away from continents for them to reach. But islands just offshore from continents (like the Channel Islands off southern California), or islands that are part of archipelagos connected in stepwise fashion to large land masses (such as Indonesia), are often close enough to allow strong swimming large mammals to establish successful island populations.
Island-dwelling animals differ from their mainland ancestors in surprisingly similar ways. The most prominent trends seen in island syndrome have already been mentioned, but some island-dwelling birds with reduced wing bones also possess enlarged bills and well-developed hind limbs. Among island mammals, pronounced changes are also observed in dentition, skeletal proportions, antler development, behaviour, and life history characteristics such as age at sexual maturity.
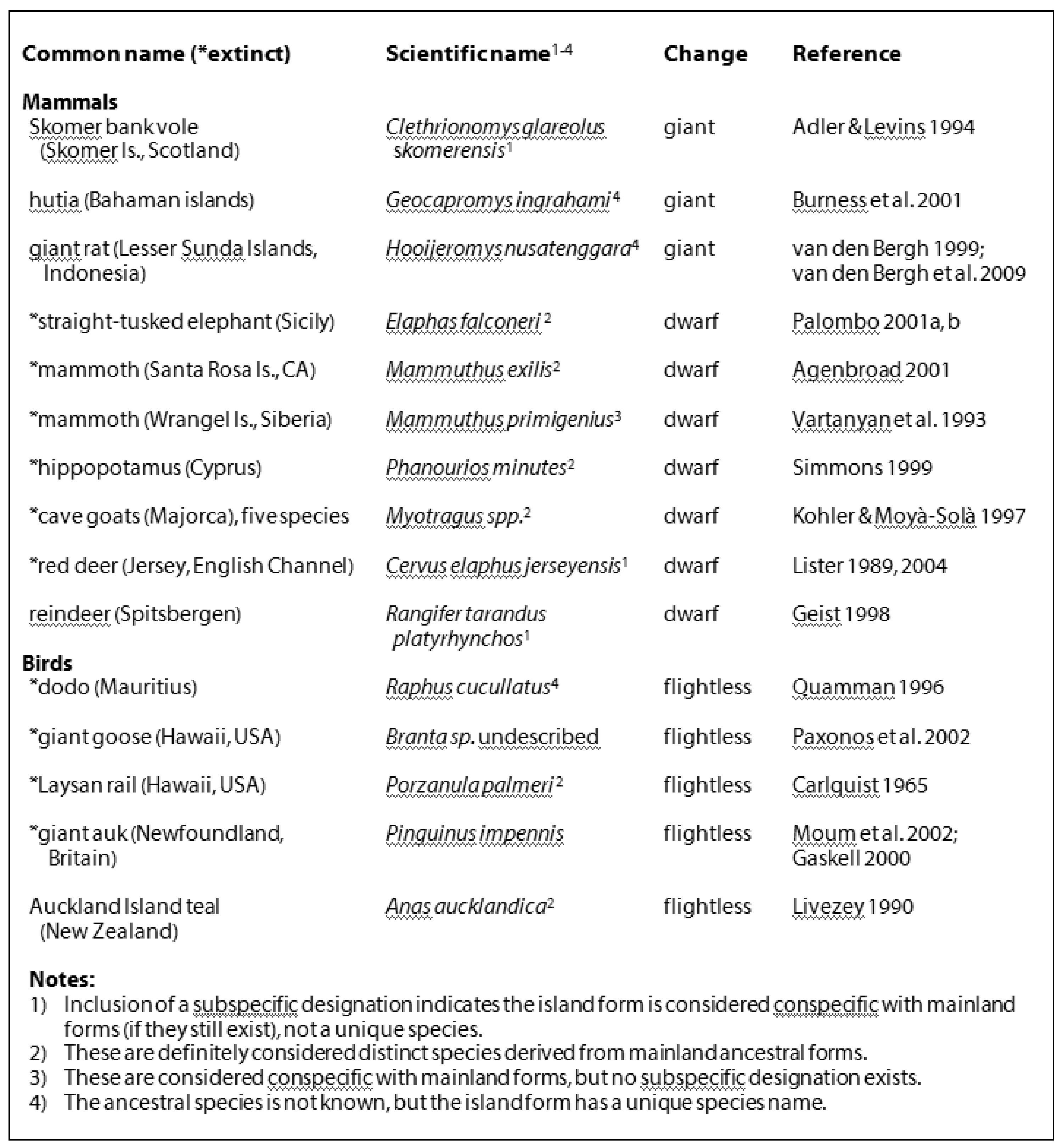
Some examples of island dwarfs and giants are listed the table below, where I’ve included a notation that emphasizes the range of variation in scientific naming among them. Some island endemics (but not others) are considered to belong to the same species as mainland forms—sometimes (but not always) distinguished by a distinct subspecies name—while others are considered distinct species (there doesn’t appear to be any rule of scientific nomenclature that governs the official naming of island forms, and some of these listed may differ from other authors consulted).
Traditional explanations for island syndrome
Regardless of how they are named, several explanations have been offered over the years to account for these trends among island-dwelling animals. These explanations typically focus on a combination of three main factors that differ between mainland and island environments: lack of competition between species, lack of predation, and either restricted or abundant food.5
A typical account suggests that a founder population of large animals will eventually become small (sometimes very small) because as their population grows, they deplete limited food resources, allowing natural selection to favour the smaller body sizes that require less food. Alternatively, founder populations of small animals (such as rodents and birds) that colonize an island often arrive to find not only abundant food but no predators or competitors for the food they require for optimal growth—in this case, the story goes, there is virtually no restriction on how large animals can become, and they grow to larger and larger sizes with each passing generation.
Some researchers believe that these selective factors alone aren’t adequate to explain the phenomenal range of body size changes that have been documented for various populations of island-dwelling animals. They suggest instead that shifts in life history traits (such as rates of mortality or the age when individuals reach sexual maturity) offer a better explanation for why island endemics change as they do.6
But while there may indeed be a significant connection between life history traits and island syndrome in a general sense, that doesn’t address the question of how the diversity of changes we see are actually implemented biologically.
In other words, I could amend my original question about dog domestication (the one that started me along this research path in the first place) and ask: what has to happen to a pigeon, in strictly biological terms, in order for it to become a dodo? Until that question is answered, we really won’t have a clear understanding of how and why islands are so often home to populations of bizarrely-shaped animals.
Parallels with domestication
Let’s deal with one kind of change at a time, starting with the most common and dramatic trend, that of size reduction, or dwarfing, of large animals. The dwarfing associated with island colonization is achieved, not by achondroplasia (a specific genetic mutation leading to disproportionately stunted growth, which occurs occasionally in all kinds of animals, including humans), but by a shift in the rates of growth. This process is known scientifically as paedomorphosis, or less formally as juvenilization, in which altered growth rates result in individuals attaining smaller sizes at sexual maturity than their ancestors.
Domestic animals, as discussed in an earlier essay here, are prime examples of this process, characterized as they are by distinctly juvenilized appearance and behaviour as compared to their wild ancestors. Indeed, for decades the same sorts of size and shape differences documented between wild ancestors and domestic descendants have been noted between island endemics and their mainland ancestors. Not only that, but many island endemics are also relatively tame or show markedly less aggression than mainland forms.7
I contend that the similarities between island-dwelling animals and domesticates are not coincidental but in fact represent evidence that an identical evolutionary process is at work. Just as the common physical and behavioural changes seen in different domestic mammals can be effectively explained as being the result of deliberate colonization of a particular habitat type by stress-tolerant individuals of wild ancestors, so can the changes seen in island species.
I maintain that animals which consciously (i.e., deliberately) colonize islands are a hormonally-similar subset of the original population—that if we were to measure thyroid hormone secretion patterns in colonizers, we would find the particular thyroid rhythms associated with stress-tolerant behaviour in those animals. As a consequence, similar trends in outcome are inevitable every time an island is deliberately colonized. Stress-intolerant animals simply choose not to participate in deliberate colonization events.’
Why choose to colonize an island in the first place? Large herbivorous mammals in particular can probably smell vegetation on offshore islands. Stress-tolerant individuals of species that are good swimmers (such as deer, elephants and hippopotamus) may occasionally cross long stretches of water to explore such offshore resources and simply decide to stay. Although colonization of islands happens under rarely occurring circumstances, once undertaken it seems not to be reversed—which means that once a few colonizers occupy an island, the ocean is as effective as a fence in keeping them there.
All new habitats are associated with novel stressors, and islands are no exception. Island colonization presents both limitations and opportunities that not only are potentially stressful but require behavioural responses:
1) Predator vigilance is still required—even if predators are not present, their absence can never be assumed.
2) Decisions are required regarding the many unoccupied habitats available for use—if no other animals are present on an island to prevent the colonizer from selecting one new habitat over another, a choice will have to be made somehow.
3) Former seasonal movements between resource areas (such as pastures around water sources that don’t dry out in the summer) are no longer possible—movement is restricted by the small physical space and oceanic boundary.
4) The small number of initial founders severely restricts mate choice for many generations.
5) Above all, there is—or appears to be—no escape.
To the initial stress-tolerant founders, these factors (predator vigilance, opportunities that require decision-making, restricted movements, limited mate choice, and virtual imprisonment) may not be especially disturbing, but they may prove very stressful for individuals born in subsequent generations who by chance inherit a thyroid rhythm that is less stress-tolerant. In contrast to domestication and other examples of colonization, on islands such stress-intolerant descendants are not free to leave. This inability to leave the new colony presents the largest difference between island colonization and domestication.
Even if there is still a swimmable distance between a continental island and the mainland, a stress-intolerant individual is as unlikely to attempt an escape from the island as to have chosen to swim there from the mainland in the first place. This is one reason I suggest that original founders must be particularly stress-tolerant, at least when colonization is by deliberate choice.
What happens when one of these less stress-tolerant second or third generation founders becomes pregnant? We know that absolute levels of thyroid hormone must rise significantly during pregnancy, often to double pre-pregnancy levels (see previous essay here), because T4 is essential for fetal development and growth at all stages.
Studies in both humans and experimental animals have shown that when T4 levels do not rise to appropriate levels during pregnancy, premature birth or reduced uterine growth are almost inevitable, resulting in smaller than usual newborns. We protect pregnant women from stress because smaller newborns face higher mortality.
Because stress of any kind reduces T4 production immediately, and often profoundly, the relationship between maternal stress and fetal growth becomes paramount to understanding island syndrome, which I’ll explain in my next essay.
The easiest thing you can do to support Biology Bites is to click the “♡ Like”. The more likes = the higher this post rises in the Substack feeds, which puts Biology Bites in front of more readers.
Crockford, S.J. (2006). Rhythms of Life: Thyroid Hormone and the Origin of Species. Trafford, Victoria.
Quammen, D. (1996). The Song of the Dodo: Island Biogeography in an Age of Extinctions. New York: Scribner.
Simmons, A.H. (1999). Faunal Extinction in an Island Society: Pygmy Hippopotamus Hunters of Cyprus. Kluwer Academic, New York.
Grant, P.R. (1998). Patterns on islands and microevolution. In P.R. Grant (ed), Evolution on Islands. Oxford: Oxford University Press. p 1-17.
Burness, G.P., Diamond, J. and Flannery, T. (2001). Dinosaurs, dragons, and dwarfs: the evolution of maximal body size. Proceedings of the National Academy of Sciences USA 98:14518-14523.
Tikhonov, A. and Vartanyan, S. (2001). Populations of wooly mammoth in North-East Siberia: dwarfing in isolation or last stage of extinction? In G. Cavarretta, P. Gioia, P. Mussi, and M. Palombo (eds), The World of Elephants: Proceedings of the First International Congress. CNR, Rome. p. 519.
Baeckens, S. and Van Damme, R. (2020). The island syndrome. Current Biology 30(8), PR338-R339. Open access https://doi.org/10.1016/j.cub.2020.03.029
Jezierski, M.T., Smith, W.J., and Clegg, S.M. (2024). The island syndrome in birds. Journal of Biogeography 51(9), 1607-1622. Open access https://doi.org/10.1111/jbi.14720
Whittaker, R.J., Fernandez-Palacios, J.M., Matthews, T.J., et al. (2017). Island biogeography: Taking the long view of nature’s laboratories. Science 357, eaam8326 https://doi.org/10.1126/science.aam8326 [not open access but has an ample summary]
Palkovacs E.P. (2003). Explaining adaptive shifts in body size on islands: a life history approach. Oikos 103, 37-44. https://doi.org/10.1034/j.1600-0706.2003.12502.x Request pdf here.
Raia, P., Barbera, C. and Conte, M. 2003. The fast life of a dwarfed giant. Evolutionary Ecology 17:293-312. https://doi.org/10.1023/A:1025577414005 pdf here.
Adler, G.H. and Levins, R. (1994). The island syndrome in rodent populations. Quarterly Review of Biology 69, 473-490.
Palombo, M.R. (2001). Endemic elephants of the Mediterranean Islands: knowledge, problems and perspectives. In G. Cavarretta, P. Gioia, P. Mussi, and M. Palombo (eds), The World of Elephants: Proceedings of the First International Congress. CNR, Rome. pg. 486-491.
Fascinating. I'd never thought about the likelihood of stress-tolerance selection because of a characteristic required to make that initial journey. Thank you!